The plant sciences provide novel strategies, new technologies, and practical solutions or mitigations to many significant challenges facing future generations.
The UVM Plant Biology Department provides research and education in the basic biology of plants at all levels of organization, from the molecule to the organism to the ecosystem. Global problems plant biologists are addressing include natural-resource sustainability, food security, global climate change, renewable energy, and human health and well-being.
UVM Plant Biology
Programs



UVM Plant Biology Undergraduate Student Opportunities



UVM Plant Biology
Facilities and Biocultural Resources



UVM Plant Biology Shared Resources
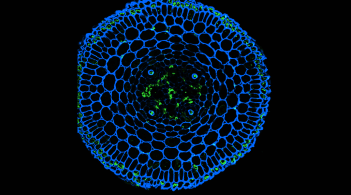


A Sweet Local Partnership
Advanced Field Ecology in the Adirondacks
Graduate Sam Fertik: Plants, Paint and the Payoff of a Multi-disciplined Perspective
Measuring Mansfield's Alpine Plants, One Square Meter at a Time
Paris Receives the 2023 President's Distinguished University Citizenship and Service Award
Announcing the FSRC PhD Fellows Class of 2023